

Около 2/1 мировых ежегодных выбросов CO2 фиксируется в органических веществах водорослями посредством фотосинтеза. По сравнению с наземными высшими растениями, фотосинтетическая секвестрация углерода микроводорослями имеет множество уникальных преимуществ: во-первых, эффективность фотосинтеза микроводорослей выше и скорость роста выше, чем у растений, во-вторых, культура микроводорослей не занимает пахотные земли и ресурсы пресной воды и может даже использовать сточные воды и очистку промышленных дымовых газов. Содержание углерода в микроводорослях близко к 50%, который в основном образуется из иммобилизованного CO2, а производство 1 кг биомассы микроводорослей (сухой вес) позволяет фиксировать около 1,8 кг CO2. Что еще более важно, производимая биомасса микроводорослей рассматривается как устойчивый источник биоэнергии и биохимических продуктов, а максимизация ценности ресурсов биомассы микроводорослей с помощью концепции биопереработки постепенно стала горячей точкой исследований во всем мире.
Биомасса микроводорослей содержит множество биологически активных веществ, имеющих лекарственную и пищевую ценность, таких как полиненасыщенные жирные кислоты, каротиноиды, витамины и т. д., а биомасса микроводорослей также может использоваться для производства различных биоэнергий в качестве заменителя ископаемой энергии, например биоводорода. , биоэтанол и биодизель. Кроме того, микроводоросли являются не только естественной приманкой для животных аквакультуры, но и служат источником высококачественного белка для питания человека, решая проблемы продовольственной безопасности.

Однако крупномасштабное выращивание и коммерческое применение микроводорослей по-прежнему сталкивается с проблемами низкой фактической урожайности и высокой стоимости выращивания. Теоретическая максимальная эффективность использования солнечной энергии микроводорослями в процессе синтеза биомассы посредством фотосинтеза составляет 8%~10%, температура листьев составляет 30 °C, а концентрация CO2 составляет 387 мл/м3, C3 и C4. Теоретические максимумы скорости преобразования солнечной энергии для растений. составляют 4,6% и 6% соответственно. Однако в большинстве случаев фактическая скорость использования световой энергии при непрерывной культуре микроводорослей в лаборатории составляет всего около 3%, а степень конверсии крупномасштабной культуры еще ниже. Это указывает на то, что культура микроводорослей еще далека от реализации своего фотосинтетического потенциала и еще есть огромные возможности для оптимизации с точки зрения фотосинтетической способности связывать углерод и потенциала технологии выращивания.
1.Оптимизация стадий фотореакций в фотосинтезе микроводорослей
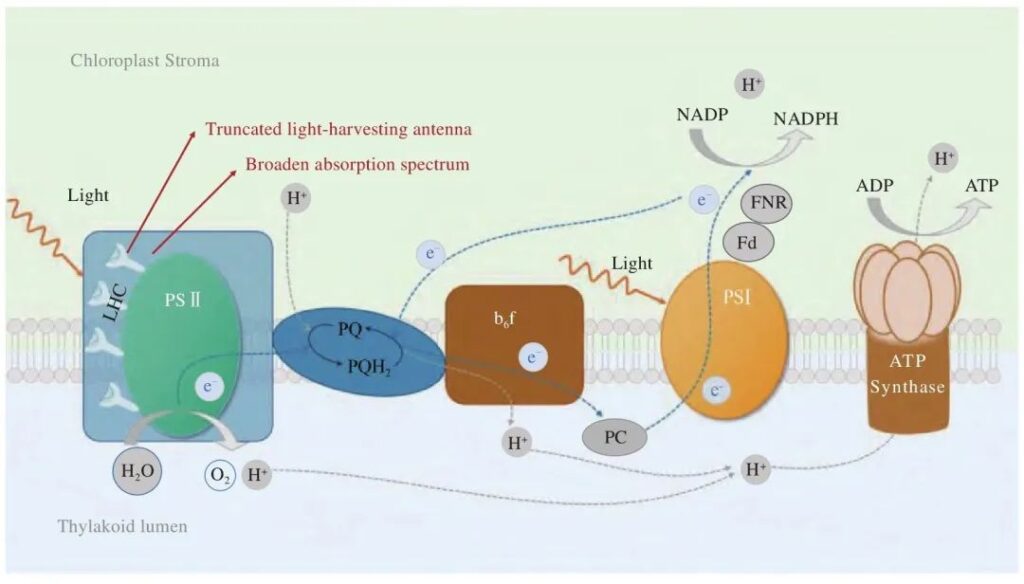
На рис. 1 представлена принципиальная схема стадий фотореакций микроводорослей. Подобно высшим растениям, микроводоросли захватывают световую энергию через различные типы светособирающих антенн (светособирающие комплексы) и проводят ее к реакционному центру фотосистемы II., где окисленная вода выделяет кислород, производит ионы водорода и электроны и в конечном итоге производит АТФ и НАДФН. Между фотосистемой II существует цепь переноса электронов. и I., который содержит акцепторы электронов, такие как пластидный хинон и цитохром b6f (Cyt b6f). Усечение светособирающей антенны и повышение эффективности использования световой энергии являются наиболее часто используемыми методами модификации стадии фотореакции микроводорослей.
2.Оптимизация темновой фазы реакции фотосинтеза микроводорослей
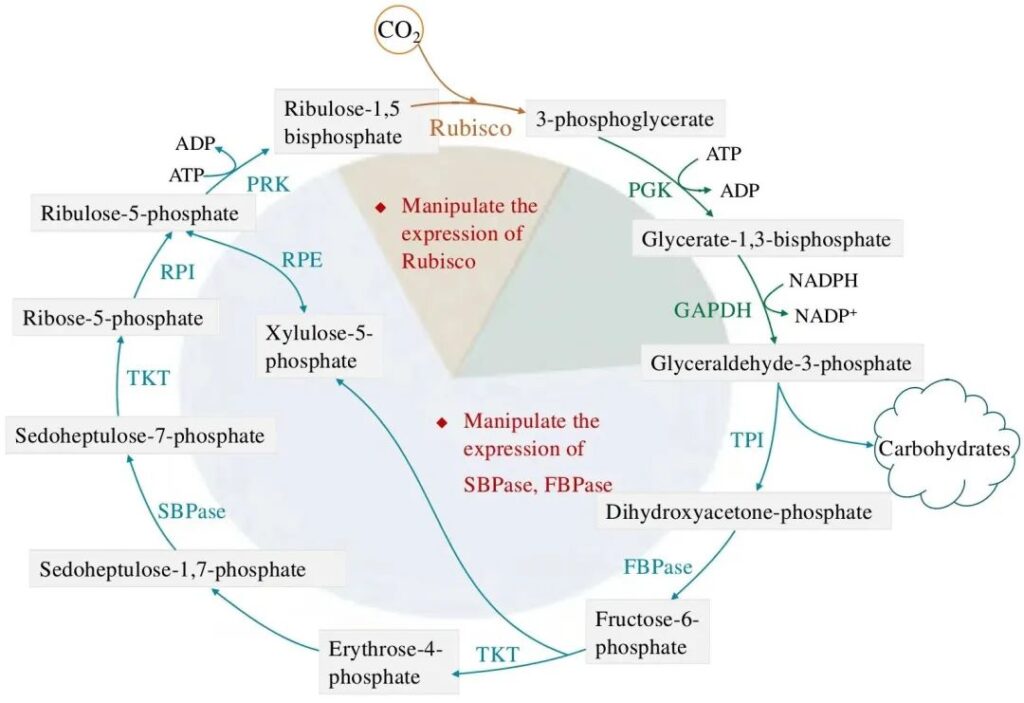
2.1 Модификация рибулозо-1,5-бисфосфаткарбоксилазы/оксигеназы
В цикле CBB рибулозо-1,5-бисфосфаткарбоксилаза/оксигеназа (Рубиско) катализирует реакцию 1 молекулы CO2 с 1 молекулой рибулозо-1,5-бисфосфата (RuBP) с образованием 2 молекул 3-фосфоглицерата, которые считается лимитирующей стадией всего цикла. Рубиско — самый распространенный белок в биосфере, образующий мост между неорганическим углеродом и живыми организмами. Но в то же время Рубиско также считается очень неэффективным катализатором. Таким образом, у высших растений Рубиско также считается основной целью фотосинтетической модификации, которая привлекла большое внимание и исследования. Строение Рубиско у цианобактерий, диатомей и зеленых водорослей очень похоже и состоит из 3 больших субъединиц и 8 малых субъединиц. Гены больших и малых субъединиц зеленых водорослей Рубиско присутствуют в хлоропластном и ядерном геномах соответственно, тогда как гены, кодирующие большие и малые субъединицы диатомей, расположены в геномах хлоропластов. Микроводоросли всегда считались отличными «донорами» Рубиско, а использование Рубиско в качестве источника микроводорослей для замены Рубиско у высших растений рассматривается как потенциальное средство повышения эффективности фотосинтеза и биомассы высших растений. Однако из-за сложного механизма сборки Рубиско гетерологичная экспрессия микроводоросли Рубиско в растениях не привела к удовлетворительным результатам.
2.2. Построение фотодыхательных ветвей.
Оксигенация Рубиско приводит к образованию токсичного метаболита 2-фосфогликолевой кислоты (2-PG), который необходимо перерабатывать по пути фотодыхания, но процесс фотодыхания требует потребления АТФ и НАДФН и высвобождения иммобилизованных CO2 и NH3; 2 молекулы 2-ПГ превращаются в 1 молекулу 3-фосфат-глицериновой кислоты (3-ФГК) посредством пути фотодыхания, и весь процесс состоит из распределения в хлоропластах, пероксисомах. Девять ферментативных стадий в митохондриях и цитоплазме завершаются, и подсчитано, что фотодыхание может высвободить до 9/1 иммобилизованного CO4, что приводит к огромным потерям C и N. Однако процесс фотодыхания восстанавливает 2% C для клеточного метаболизма из 2 молекул 2-PG, который играет очень важную роль. играют важную роль в клеточном метаболизме, поэтому теоретически построение новой ветви фотодыхания, чтобы избежать потерь CO75 и NH2 или увеличить способность восстановления высвобождения CO3, может улучшить CO2 и увеличить скорость производства биомассы. Реализуемость этой схемы была успешно продемонстрирована на модельном организме Arabidopsis thaliana и рыжике масличном, а вновь построенная фотодыхательная ветвь может помочь трансформированному штамму значительно увеличить скорость производства биомассы.
2.3 Трансформация механизма обогащения углерода
В отличие от растений C3, большинство микроводорослей и растений C4 имеют механизм концентрации углерода (CCM), позволяющий справляться с более низкими концентрациями CO2 в водной среде. У цианобактерий Рубиско запечатан в карбоксисомах, тогда как у эукариотических микроводорослей Рубиско в основном присутствует в белковых ядрах хлоропластов, а концентрация CO2 вокруг активного центра Рубиско может более чем в 1000 раз превышать концентрацию CO2 в окружающей среде благодаря CCM. Карбоангидраза (КА) является ключевым ферментом, участвующим в CCM, и отвечает за катализацию обратимого взаимодействия между HCO3- и CO2. CCM в микроводорослях обычно представляет собой индуцируемый механизм, который регулирует уровень экспрессии CCM посредством восприятия окружающих концентраций CO2. При крупномасштабном выращивании микроводорослей часто используются концентрации CO2 значительно выше атмосферных, что может привести к отключению СКК.
Таким образом, многие важные новаторские успехи были достигнуты в трансформации и оптимизации фотосинтетических систем микроводорослей посредством усечения светособирающей антенны, увеличения светособирающей способности и использования световой энергии, преобразования цикла CBB и CCM, а также создания фотодыхательная ветвь. Однако общий прогресс отстает от трансформации и оптимизации путей фотосинтеза высших растений, особенно товарных культур, что приводит к нереализованному потенциалу связывания углерода микроводорослями. Благодаря добыче превосходных фотосинтетических элементов, регуляторных факторов и созданию новых путей можно эффективно снять ограничение на узкие места и реализовать энергетический баланс между световыми и темновыми реакциями. Фотосинтез не только регулируется светом, но и регулируется эффективностью потребления конечного продукта, что указывает на необходимость одновременного осуществления модификации метаболических путей, связанных с фотосинтезом, на клеточном уровне. Благодаря быстрому развитию синтетической биологии была заложена основа для разработки и создания высокоэффективных инженерных штаммов микроводорослей, улавливающих углерод, с использованием микроводорослей в качестве организмов-шасси для фотосинтетической секвестрации углерода. Методы и концепции синтетической биологии могут использоваться для разработки или синтеза фотосинтетических систем с более высокой эффективностью световой энергии и эффективностью связывания углерода, устойчивостью к фотоповреждениям и производством менее активных форм кислорода, независимо от видовых ограничений.
О БайЛун Биотехнологическая Компания, ООО:
БайЛун Biotechnology Co., Ltd. является ведущим поставщиком и ведущим поставщиком технических услуг, специализирующимся на поставке комплексных биореакторных систем и передовых решений по управлению. Наша обширная линейка продукции включает в себя широкий спектр предложений: от биореакторов (ферментаторов) до биореакторов на животных клетках, биологических шейкеров и систем управления, специально разработанных для биотехнологических процессов. Имея емкость от 0,1 л до 1000 л, мы стремимся способствовать росту китайской биореакторной промышленности в глобальном масштабе.
В БайЛун, мы можем похвастаться опытной командой инженеров, обладающих глубокими знаниями в области процессов ферментации, биохимического оборудования и химических технологий. Более того, мы активно привлекаем всемирно известных экспертов и ученых в качестве технических консультантов, обеспечивая надежную технологическую основу нашей продукции. Центральное место в нашем духе занимает неустанное стремление к инновациям в продукции и технологическому лидерству, и все это направлено на обеспечение максимальной удовлетворенности клиентов. Мы считаем себя ответственными за приоритетность преимуществ для клиентов, воплощая это обязательство в качестве краеугольного камня основных ценностей компании Bailun.
Наш разнообразный портфель продуктов тщательно разработан для удовлетворения разнообразных потребностей наших клиентов. С самого начала мы придерживались принципов приоритета качества, клиентоориентированности и честности операций. Наша непоколебимая приверженность удовлетворению растущих потребностей наших клиентов двигает нас вперед. Принимая волну экономической глобализации, мы искренне приглашаем к сотрудничеству предприятия по всему миру, стремясь к взаимному процветанию и успеху.
Дух БайЛун воплощает в себе приверженность клиентоориентированности, стремление к совершенству в области качества, приверженность справедливости и порядочности, а также непоколебимое стремление к постоянному совершенствованию и инновациям.
Связаться с нами:
Добавить: 6848# Liuxiang Rd., Цзядин, Шанхай, Китай
Контактное лицо: Мэдди
Email:sales@fermentertech.com
Телефон:+86-134-7276-8163 (WhatsApp)
Веб-сайт: https://fermentorchina.com/

